PhD Project
Insights into Cenozoic fishes (Chondrichthyes, Teleostei) from South America and Europe
PhD Student

Dr. Jaime A. Villafaña Navea
ORCID: 0000-0002-6441-9025
ResearchGate
Supervisors
- Univ.-Prof. Dr. J. Kriwet (University of Vienna, Austria) [supervisor]
- Marcelo M. Rivadeneira (Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Coquimbo, Chile) [co-supervisor]
Grant
Becas-Chile Scholarship (Becas para estudios de doctorado en el extranjero, National Research and Development Agency of Chile)
[ad personam]
Aims
The overarching goals of this project were to (1) provide new insights into the faunal composition and diversity of Cenozoic fishes from central Europe and the eastern Pacific of South America and (2) analyse the biogeographic dynamic and diversity patterns of central Europe and South American fish assemblages. The specific aims were to improve and complement the previous knowledge of (1) eagle rays, horn sharks and sawsharks from the Neogene of the eastern Pacific of South America; (2) chondrichthyans from the early Miocene of Chile and Oligocene/early Miocene boundary of Ecuador; (3) elasmobranchs (sharks, rays, skates) from the western Paratethys of Germany and their relation with other faunas from Europe; (4) diversification patterns (richness, origination and extinction rates) of European chondrichthyans and their relation to tectonic, oceanographic and climatic events that occurred during the Neogene on regional and global scales. In addition, the high abundance of fossil remains of the great white shark Carcharodon carcharias from the eastern Pacific was analyzed from an ecological point of view.
Conclusions
Most of the studies of the fossil record of chondrichthyans from the eastern Pacific of South America are focused on fossiliferous localities ranging from the middle Miocene to Pliocene (Carrillo-Briceño et al., 2018; Villafaña and Rivadeneira, 2018, and references therein). Therefore, the faunal composition and distribution of this group from older time intervals (i.e., late Oligocene-early Miocene) of the eastern Pacific of South America is poorly known and understood. In order to cover this lack of information, Jaime collaborated in a publication about the Oligocene-early Miocene fossil record of chondrichthyans from Ecuador. Of the total described taxa (2 orders, 1 family, 8 genera, and 16 species), 19 represent new records for Ecuador, 10 new records for the Central Eastern Pacific and four new records for South America. Two paleobathymetric analyses were performed using the described taxa to estimate the most probable depositional environment, which was determined as outer neritic.
In another publication, Jaime reviewed and described the fossil record of early Miocene chondrichthyans from Chile. In this study I was able to describe 8 new records (1 superfamily, 1 family, 2 genera, and 4 species) from the early Miocene of Chile, including a new extinct species of sawshark Pristiophorus homboldti. All the previous and new findings described in this publication indicate that: 1) the faunal composition of early Miocene chondrichthyans from Chile was different in comparison to the rest of the Neogene and also to that reported from other localities along the eastern Pacific, and 2) the fauna from the early Miocene of Chile experienced significant biogeographic changes from the past to the recent.
As was stated above, the fossil record of chondrichthyans from the middle Miocene to Pliocene of the eastern Pacific of South America was widely described in previous publications, however, some groups were less studied than others. For sharks, the genus Heterodontus forms part of this group, being only reported from some localities along the Chilean and Peruvian coasts (Muizon and DeVries, 1985; Carrillo-Briceño et al., 2013). Jaime collaborated as a second author in a publication that described in detail for the first time the tooth morphology of this heterodontid genus from the eastern Pacific of South America. The morphological characteristics of both lateral anterior teeth described in this study agree with those of the Francisci-type (Reif, 1976).
Conversely to Heterodontus, the great white shark (GWS) is the second most abundant species in the temperate Pacific of South America (Villafaña and Rivadeneira, 2018). He analysed the fossil record of the GWS from three Pliocene localities (Caldera, Coquimbo and Pisco) based on the main criteria to define paleo-nursery areas (high frequency of juveniles, food availability, and shallow-water depths). According to the size and life stages established for extant specimens (Fergusson, 1996; Boldrocchi et al., 2017), the specimens from Caldera (Chile) and Pisco (Peru) mostly represent subadults (≥300 to 360 cm for males and ≥300 to 450 for females), whereas juveniles (175 cm to 300 cm) dominate in Coquimbo (Chile). Therefore, the results of this publication suggest that Coquimbo is the first nursery area of the GWS in the fossil record.
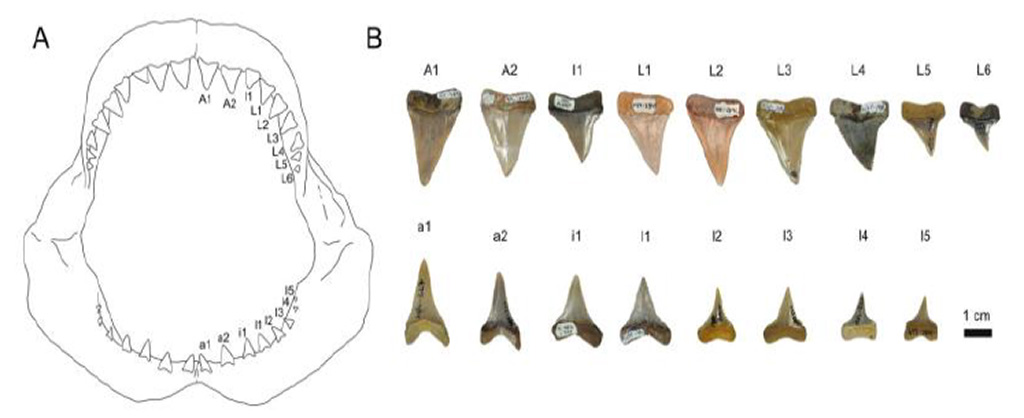
In addition to the new species Pristiophorus humboldti, another sawshark genus was recently described from the middle Miocene-early Pliocene of Chile by Jaime. In this study, he collaborated in the first description of the genus Pliotrema, which only was mentioned in previous studies but not studied in detail (Case, 1973; Welton, 1974). The specimen described in this publication represents a well-preserved rostral spine, which shares the morphological characters for its assignment to the genus Pliotrema. However, the diagnostic features used to identify it on generic level (i.e., serrations along the posterior cutting edge) is distinctly different to that observed in any known extinct and extant specimens.
In the case of batoids from the eastern Pacific of South America, Jaime described in detail the fossil remains of the genus Aetomylaeus from one locality in Peru and six localities in Chile. The material described can be unambiguously assigned to genus Aetomylaeus based on the morphological characters observed in extinct and extant specimens within the genus. Also, this study confirms the high variability of tooth row numbers and the morphological variation within tooth rows in the genus Aetomylaeus (Hovestadt and Hovestadt-Euler, 2013).
Although the eastern Pacific of South America hosts the richest fossiliferous localities in South America with marine vertebrate remains (including sharks), records of bony fishes are poorly known (e.g., Long, 1993; Walsh, 2001). For this, he collaborated in a study, which will increase the knowledge of the bony fish fossil record along the eastern Pacific of South America. In this study, the team described 27 fossil remains, including dentaries, neurocrania, pharyngeal tooth plates, opercular bones and rostra. The fauna includes the first records from Chile (Sardinops cf. sagax), the eastern Pacific of South America (e.g., Labrodon sp.), South America (Sarda sp.), the Southern Hemisphere (e.g., Semicossyphus sp.) and the Neogene period (Gymnosarda sp.). All this assemblage contributes to understand the past diversity of fishes in Chile and its relation with other localities along the temperate Pacific of South America. In addition to allowing us to infer the depositional paleoenvironments of this geological formation based on a paleobathymetric analysis.
In this project, Jaime also considered Europe as a study region of the chondrichthyan fossil record. He described an elasmobranch assemblage from the lower Miocene Upper Marine Molasse of the Simsee area in Southern Germany. The 37 elasmobranch taxa reported include 26 sharks and 11 batoids. The biogeographic analysis of the Simsee fauna from the past to the recent show different biogeographic dynamics. These changes could have been influenced by the intense tectonic, climatic and oceanographic events that occurred during the early Miocene of Europe (Rögl, 1999). The results of this study confirm the similarity of closely adjacent localities in Europe due to the connection through seaway passages.
In a final study, he analysed the biogeographic dynamics of the studied associations, which are complex with 27% of the genera from the Neogene of Europe being now globally extinct whereas the proportion of regionally extinct genera varied (40-56%) depending on the analysed region. The result of the diversity trajectories also shows marked differences between regions. In general, the peaks in diversity (i.e., high number of occurrences, genera and origination rates) are mainly associated to warm climatic events and the opening of sea passages between the regions during the early and middle Miocene (Nebelsick, 1992; Rögl, 1999). In the case of the extinction rates, the peaks also coincide with regional and global climatic (i.e., cooling episodes) and oceanographic events (Gürs, 2001; Westerhold et al., 2020). (For references see Jaime’s publications and PhD thesis.)
PhD Theis
The resulting PhD thesis summarised all new findings of European and South American fishes during the Cenozoic. In South America, the knowledge of the diversity and biogeographic dynamics of chondrichthyans from older intervals i.e., Oligocene-early Miocene (Chapter 1, Papers 1-2) and less studied groups i.e., horn sharks (Chapter 1, Paper 3), sawsharks (Chapter 1, Paper 5) and eagle rays (Chapter 1, Paper 6) was increased. New ecological interpretations explain the high abundance of the great white shark in the temperate Pacific of South America, i.e., nursery area (Chapter 1, Paper 4). The fossil record of bony fishes from the region was also studied, showing a high diversity of these groups during the late Neogene of Chile (Chapter 1, Paper 7). In Europe, the rich cartilaginous fauna described from southern Germany displays different biogeographic dynamics (Chapter 2, Paper 8), possibly explained by the intense climatic and oceanographic events during the early Miocene. The relation between the biogeographic and diversity dynamics of Neogene chondrichthyans from Europe with the climatic and oceanographic events is a current work in progress (Chapter 3, Paper 9).
Future research directions
Although the knowledge of the fossil record of South American and Central Europe faunas was increased during this dissertation, descriptive and analytical studies are still needed. Some of the possible future research directions of each publication described in this PhD thesis are discussed in following paragraphs.
The chondricthyan assemblage reported from the fossiliferous localities of the Montañita-Olón outcrops is the most diverse known from Ecuador and for the late Oligocene-early Miocene of Tropical America (Chapter 1, Paper 1). However, new findings could help to improve the indentifications. For instance, seven taxa were described at genus level (e.g., Dalatias) while eight to unconfirmed species (e.g., Centrophorus cf. granulosus). Additionally, the confirmed presence of the species used in the paleobathymetric and habitat preference analysis (e.g., will contribute to refine the interpretations of the most probable depositional environment of the Dos Bocas Formation.
Another chondrichthyan assemblage from the early Miocene was dscribed from Chile (Chapter 1, Paper 2). This material was collected by Dr. Sven Nielsen (Universidad Austral de Chile) during fieldworks in Chile in the years 2000-2002, and subseqauently was deposited at University of Vienna for examination by me in the years 2017-2018. All the specimens finally were sent back to Chile and deposited at the Chilean National Museum of Natural History. The presence of this collection in Chile will contribute enormously to research in comparing this published material with new material, collected in future campaigns. Additionally, new collected material could contribute to describe specimens at lower taxonomic levels which were poor represented in the published collection. The description of new genera and species currently inhabiting warm waters may help us to confirm the previous hypothesis that the early Miocene of Chile was mostly dominated by tropical faunas in comparison to the rest of the Neogene (Nielsen and Glodny, 2009).
The assignment of Heterodontus teeth collected from Chile to the Francisci-type (Chapter 1, Paper 3), provides more detail about the most probable identification of the genus during the Neogene, separating it from the Portusjacksoni-type. In order to identify the specimens on species level, future studies should be focused on the comparison of the tooth morphology (using geometric morphometrics) of species within the Francisci-type from the eastern Pacific (H. francisci, H. quoyi, and H. Mexicanus). The identification on lower taxonomic levels could help us to know whether the Heterodontus teeth from Chille correspond to an extinct species or if it survided and migrated to other regions as it is postulated for other sharks (Cione et al., 2007).
The study about the first nursery area of the GWS (Chapter 1, Paper 4) opens the discussion whether this nursery-area was unique in the world during the Pliocene or whether others were also present in other regions. Furthermore, other aspects about the ecology and evolution of the GWS could be included in future studies. For instance, sea surface temperature reconstructions suggest warmer conditions than today until about 3 Ma (Dekens et al., 2007; Dowsett et al., 2013) and decreasing afterwards towards colder conditions. Therefore, the relation of the decrease of temperature from the past to the recent in the region and aspects as the body size evolution of the GWS (e.g., increase of size over time) could be included in future analysis.
The confirmed presence of Pliotrema from Chile (Chapter 1, Paper 5) gives us more information about its past distribution in the eastern Pacific of South America. However, future studies based on rostral spines should consider the serration types in their descriptions, as they could contribute to knowing whether the morphological characters of the specimen from Chile are also present in records from other localities, and whether this trait potentially defines a new, hitherto unknown species. In addition, samples of microteeth need to be studied with a microscope in order to identify small-sized taxa, which has not been done up to now.
In the case of the fossil remains of the genus Aetomylaeus (Chapter 1, Paper 6) from the eastern Pacific of South America, identification at species level would be possible if better preserved fossil specimens are analyzed in addition to the study of the dental diagnostic characters of all the current species and their ontogenetic variation (Hovestadt and Hovestadt-Euler, 2013). Future studies are highly necessary to reconstruct the evolutionary history and biogeographic dynamic of this eagle ray from the Neogene to the recent in the temperate Pacific of South America.
While the fossil record of bony fishes from the eastern Pacific of South America is increased (Chapter 1, paper 7), the knowledge about their past diversity and distribution in the region is still insufficient. Future studies should focus on collecting in situ material with detailed geological information of each fossiliferous locality. In addition, the taxonomic resolution for some specimens (e.g., neurocrania) could be improved wether the material is better cleaned. Geometric morphometric methodologies could be also used to compare fossil and extant species (Brambilla et al., 2021).
The results of the faunal similarities between localities in Euope (Chapter 2, Paper 8) give us the possibility to analyze the biogeographic dynamics of elasmobranch assemblage during the early Miocene. Future studies should be focused on the modulating role of ecological and life-history traits on the biogeographic dynamics of elasmobranchs from European localities (Villafaña and Rivadenira, 2018). In addition, the study of less known localities from Europe (e.g., southern Spain) could contribute to improve the interpretation.
The preliminary results of the study in preparation (Chapter 3, Paper 9) provide the first steps to synthesize and understand the macroevolutionary history of chondrichthyans from the Neogene of Europe. However, the ages of some of the localities should be revised and uptaded based on the most recent studies (e.g., Kováč et al., 2018; Sant et al., 2020). In addition, other methodological approaches as CMR (Liow and Finerelli, 2014; Payne and Heim, 2020) and Bayesian methods (Pimiento et al., 2017; Silvestro et al., 2019) should be tested to confirm the consistency of these results. Future research studies could analyse the role of the ecological and life-history traits of each genus in the diversity patterns (richness, orgination and extinction) through time.