|
|
|
|
|
|
 |
|
Ecological effects of competition on pycnodont fishes (Actinopterygii, Neopterygii, Pycnodontomorpha) and its possible role leading to their extinction |
(FWF-Project P 29796-B29) |
Primary Investigator: Univ.-Prof. Dr. Jürgen Kriwet
Team
- Univ.-Prof. Dr. Jürgen Kriwet (PI)
- John J. Cawley, MSc (Predoctoral Researcher)
- Fabrizio De Rossi (Student Assistant)
Collaborators
- Prof. Dr. Giorgio Carnevale, Università degli Studi di Torino, Italy
- Dr. G. Marramà, Università degli Studi di Torino, Italy
Contents
- Summary
- 1. Introduction
- 2. Success of pycnodontiforms
- 3. Goals
- 4. Specific subjects
- 5. Innovative aspects, expected outcomes, and intellectual merit
- 6. Results
- 6.1 Taxonomy
- 6.2 Sexual dimorphism
- 6.3 Evolutionary success and extinction
- 6.4 Intrarelationships, compltness of fossil specimens and fossil record
- 7. Publications
- 8. Conference abstracts
- 9. Conference participations
|
|
|
|
Summary |
Pycnodonts are an ancient and iconic group of extinct fishes. Although extinct today, they were extremely successful for almost 175 million years originating ca. 227 million years ago and surviving the extinction events at the Triassic/Jurassic (ca. 200 million years) and the Cretaceous/Palaeogene (ca. 65 million years) boundaries. In the early Eocene (ca. 52 million years) they finally disappeared without any living relatives. During the Mesozoic, they rapidly diversified in both species numbers and morphological bauplans and are generally considered the coral fishes of this period occupying fundamental and important ecological niches in many shallow marine ecosystems. The reasons for their seemingly rapid diversification early in their evolutionary history, subsequent success and thus their possible superiority over other fish groups inhabiting same environments, habitat use through specific adaptations, possible size aggregations due to ontogenetic shifts, reasons for diversity fluctuations, and events triggering their final disappearance more or less simultaneously with the appearance and evolution of teleostean fishes, which are major elements of modern coral-fish assemblages still are ambiguous or have not yet been studied with rigorous analytical methods. We intend to determine possible correlations between diversification patterns and biotic factors, particularly competition, that enabled pycnodonts to occupy their niches despite supposed competition by ecological similar fishes during their evolution, and whether competition from teleosts in the Eocene triggered their extinction. To answer these questions, a multidisciplinary palaeobiological approach is necessary that will allow us to explore morphological changes, adaptations, and ecological functions of pycnodonts, and to subsequently identify possible competition patterns within pycnodonts and between pycnodonts and other fishes with similar adaptations. For this, we will analyse their phylogenetic relationships, the quality of their fossil record for ascertaining the reliability of data for studying their evolutionary history, and reconstruct their ecological role based on morphospace analyses within the habitats they occupied through time. By doing this, we also will identify possible body size segregations, which might be caused by intra- and inter competition patterns and to understand the relationships between morphological changes and increases in species numbers through time (chronophylomorphospace). This project thus will significantly augment our understanding of macro- and co-evolutionary patterns of fishes through time by integrating different aspects. We expect that the results will significantly advance and inform how certain groups prevail over others and give insights into the evolutionary success and ecology, but also into causes triggering extinctions of marine vertebrates. Innovative aspects of this project are the integrated approaches and analytical procedures, combining up-to-date methodologies.
|
up |
|
|
1.Introduction |
Pycnodont fishes (Pycnodontiformes) are a monophyletic and ecologically successful clade of extinct ray finned fishes with a fossil record spanning nearly 180 million years from the Late Triassic to middle Eocene (Tintori 1981; Poyato-Ariza & Wenz 2002; Kriwet & Schmitz 2005; Voss et al. 2019). They are most commonly associated with shallow water marine habitats but are also found in association with freshwater and estuarine deposits (Longbottom 1984; Poyato-Ariza et al. 1998). A deep, rounded, and laterally compressed body, a frontal flexure of the skull in profile view, and elongated dorsal and anal fins, forming together with the caudal fin an effective rudder characterize pycnodonts (Fig. 1). In their body shape they superficially resemble extant coral reef fishes like butterflyfishes (Chaetodontidae), surgeonfishes (Acanthuridae), and triggerfishes (Balistidae) (Fig. 2). Distinctive characteristics of pycnodonts are their molariform teeth, which generally are arranged in well-defined rows on the unpaired vomer (upper jaw) and paired prearticulars (lower jaws). This dentition represents an adaptation to shelled prey. Their diversity is strikingly high, with over >600 nominal species in >80 genera of which many taxa are based on teeth and dentitions only (Kriwet, 2001a; 2005). The palaeobiogeography of these fishes suggests they originated in the Tethys Sea initially and then spread worldwide (Kriwet 2001b; Martín-Abad & Poyato-Ariza 2013).

| Fig. 1.Examples of pycnodontiform fishes. (a) †Brembodus ridens (MCSNB 4901); (b) †Proscinetes elegans (BSPG AS I 1213); (c) †Gyrodus hexagonus (BSPG 1989 XII 110); (d) †Gebrayelichthys uyenoi (CLC S-538); (e) †Ichthyoceros spinosus (MNHN HAK 106); (f) †Corusichthys megacephalus (CLC S-596); (g) †Gladiopycnodus karami (CLC S-393); (h) †Arduafrons prominoris (NHMUK P.8658);
(i) †Akromystax tilmachiton (MNHN NRA 95); (j) †Piranhamesodon pinnatomus (JME-ETT4103). [modified from Cawley et al. (2021)]
| Traditionally considered an order, Pycnodontiformes (Poyato-Ariza & Wenz 2002), Nursall (2010) assumed pycnodont fishes to constitute a superorder Pycnodontomorpha, which was considered as a possible sister group candidate to Teleostei. However, a more recent analysis by Poyato-Ariza (2015) identified pycnodonts to be the sister group to Halecostomi (Holostei + Teleosteomorpha), which would make pycnodonts the most basal group among neopterygian fishes. Such a result indicates that the shared characters of pynodonts and teleosts are the result of convergence, not homology. Since pycnodonts seemingly no longer represent the sister group to Teleosteomorpha (stem teleosts and Teleostei) which was proposed by Nursall (2010) and all pycnodonts now belong to a single order Pycnodontiformes, and Pycnodontomorpha is no longer a valid clade. Pycnodontiformes comprises eight known families considered valid. These are, in alphabetical order: Brembodontidae, Coccodontidae, Gebrayelichthyidae, Gladiopycnodontidae, Gyrodontidae, Mesturidae. Pycnodontidae and Serrasalmimidae. In this dissertation whenever the term pycnodont is used, it refers to the total group Pycnodontiformes.

| Fig. 2.Reconstruction of †Flagellipinna rhomboides Cawley & Kriwet, 2019 from the Upper Cretaceous (Cenomanian) of Lebanon. [Arrt work by Fabrizio De Rossi]
| One of the defining characteristics of pycnodonts and indeed a major factor in their evolutionary success is the arrangement of the jaws which are unlike that of any other actinopterygian (Kriwet 2001c). The premaxilla and dentalosplenial are elongate, thin bones with a loose attachment to the skull with typically incisiform teeth for grasping prey. The main tooth bearing bones are the singular vomer (upper jaw) and the paired prearticulars (lower jaw) (Fig. 3). A pavement of molariform teeth lies on these bones, which grind against each other to process their prey. The prearticular bones form a medial trough with the lateral teeth of said bone occluding strongly against the vomer. A high coronoid process not only suggests a high bite force to deal with tough food items such as mollusks and echinoderms but the posterior position of the process also indicates fine, precise movements of the lower jaw which indicates a sort of oral chewing motion more similar to that seen in mammals than other actinopterygians (Baines 2010). Tooth microwear analysis (Baines 2010, Darras 2012) and the presence of a symphysis between the prearticulars (Gosline 1965) are further evidence of chewing behaviour. The work by Baines (2010) in particular, indicates that pycnodont jaws would have moved in a propalinal motion with the facets of teeth shearing against each other. Changes in microwear texture orientation indicate rotational movements of the prearticulars upwards and forwards to allow occlusion between prearticular and vomerine dentition.
One of the most distinctive aspects of the suspensory apparatus in pycnodonts is the reduced number of branchiostegal rays. While low numbers of branchiostegal rays can also be found in primitive actinopterygian lineages such as haplolepids, saurichthyids, redfieldiiforms and lepisosteiforms (Lambers 1991) and some teleosts (McAllister 1968), the lowest number is typically three. In contrast, the usual number of branchiostegal rays in pycnodonts is two acinaciform (slender as classified by McAllister 1968) type rays with the highest number being three (Poyato-Ariza and Wenz 2002; Ebert 2016). This reduction in number and size indicates a small branchiostegal membrane and would not permit much expansion of the opercular chamber (Kriwet 2001c). This would severely limit the degree to which pycnodonts could utilise suction feeding and as a result, biting and manipulation were the dominant mode of primary feeding strategies that pycnodonts were adapted for (Poyato-Ariza 2005a).
While typically seen as strict durophages, this range of jaw mobility and tooth variability hints that some taxa of pycnodonts could be omnivorous generalists (Poyato-Ariza 2005a) or even herbivores (Vullo et al. 2019). The recent discovery of short jaws with blade-like teeth from the Palaeocene belonging to a family of piranha mimicking pycnodonts with the appropriate name of Serrasalmimidae, shows that even piscivory could be well within the bounds of what pycnodont jaws were capable of doing (Vullo et al. 2017). This is further confirmed by the discovery of Pirahnamesodon pinnatomus from the Plattenkalk deposits of the Late Jurassic (Late Kimmeridgian - Early Tithonian) of Ettling (Kölbl-Ebert et al. 2018). This shows that the image of pycnodonts as strictly shell crushers is becoming increasingly inaccurate.

| Fig. 3.Right prearticular dentition of †Gyrodus sp. (NLH 102.971) from the lower Barremian of clay pit “Gott” in occlusal view. [Modified from Kriwet & Schmitz 2005]
In terms of morphology, pycnodonts are usually laterally compressed, deep-bodied fishes (Poyato-Ariza and Wenz 2002), which is similar to modern reef fishes such as Pomacanthidae and Chaetodontidae. The elongated dorsal and anal fins of many pycnodonts are similar to fishes from the family Balistidae, which swim by simultaneously undulating the unpaired fins (Sfakiotakis et al. 1999), which makes pycnodonts typically slow but manouverable. Pycnodont species show great variation in medial and caudal fin morphology (Poyato-Ariza 2005a). Gyrodus have falcate dorsal and anal fins along with a forked caudal fin, which together with their wide distribution hints at this fish being more pelagic than other pycnodonts (Kriwet and Schmitz 2005) and capable of long distance movements. The narrow caudal peduncle and strip-like medial fins of Pycnodus (Blot 1987; Cawley et al. 2018) also indicated adaptations for more sustained swimming in comparatively open waters in contrast to more ‘typical’ pycnodonts. Recently discovered pycnodont families from the carbonate reef platform of the Late Cretaceous of Lebanon reveal a far deeper variety of pycnodont morphologies than previously thought. Gladiopycnodontidae with their elongate bodies, large heads and assortment of spines suggest a more defensive, pelagic mode of life (Taverne and Capasso 2013; Marramà et al. 2016a). Even stranger are the Gebrayelichthyidae, where their bodies are extremely antero-posteriorly shortened yet their high skull roof and low pectoral girdle contribute to one of the highest bodied actinopterygians known (Nursall and Capasso 2004). This unusual body shape along with reduced fins suggests a very slow swimming fish with high maneuverability.
A common characteristic of pycnodonts is the autogenous anterior neural spines. These are neural spines, which have separated from the basidorsal of the ossified notochord. Nursall (1996) suggests that the purpose of these spines is to allow for extra kinesis in the cranium, which would have enabled pycnodonts to increase the force of their bites and jaw gape. It is a generally observed trend that more plesiomorphic taxa (e.g. Brembodus with most neural spines including those in the caudal region (Tintori 1981; Kriwet 2001a; Poyato-Ariza and Wenz 2002)) have a much higher number of autogenous spines than more derived pycnodonts (e.g. Abdobalistum) with some having less than six. This high number of spines not attached to the vertebral column along with absent ossified vertebral centra lead to anteriorly directed forces not being rapidly generated and would lead to these forms having a poor fast-start performance. The pycnodontiform Gyrodus is unusual as despite having a high number of autogenous spines (14), it also possesses a well-developed caudal peduncle that would allow for increased swimming performance and speed (Kriwet and Schmitz 2005).
The above adaptations were most likely significant contributions to the evolutionary success of pycnodonts and would have enabled them to weather both the Late Triassic/Early Jurassic and the K/Pg extinction events. Indeed, their increasing raw species diversity through the Mesozoic is quite plainly seen in the fossil record (Kriwet 2001a). The increase in raw species numbers in the Late Cretaceous is mirrored in the great increase in morphological disparity (Marramà et al. 2016a). The increase in species diversity from the Middle Jurassic onwards indicates the first diversification event in the history of pycnodonts.
A steep drop in taxon number occurred from the Late Cretaceous to Palaeocene, which might indicate that the K/P boundary event also affected pycnodont fishes. The disappearance of most major lineages well before the K/P boundary, which was used by Kriwet (2001a) to reject an extinction event at this boundary might be related to the so- called Signor-Lipps effect.
A popular hypothesis regarding the extinction of pycnodonts is the observation that many modern teleost groups common in coral reefs today arose at the same time that pycnodonts were beginning to decline (Bellwood 1996; 2003; Poyato-Ariza 2005a). Were the pycnodonts simply outcompeted by better-adapted teleosts or could there have been other factors in their extinction?
During the Mesozoic, pycnodonts also faced competition from shallow water adapted, shell crushing ginglymodian and dapediiform fishes. In this case, however, the ginglymodian fishes (with the exception of Lepisosteidae) perished at the K/Pg boundary (Cavin 2010) while the dapediiforms did not survive past the Kimmeridgian-Tithonian of the Late Jurassic (Lambers 1999; Woodward 1895). Are there correlated evolutionary trajectories in all three groups that enabled some to survive and others to perish? It is these questions that this dissertation will primarily address.
|
up |
|
|
2.Success of pycnodontiforms |
Many scientists reflected on the ecology of pycnodonts and also their possible success. Their laterally compressed, high, rounded bodies and long dorsal and anal fins show a striking resemblance to modern coral-fishes (Poyato-Ariza 2005). This morphology is highly suited to shallow water marine reefs and pycnodont fossils are usually used as indicators of such a habitat wherever their fossils are discovered. However, the interpretation that pycnodonts were exclusively reef dwellers is mistaken, because many modern teleost groups thought to be exclusive reef dwellers such as Acanthuridae are also found in a variety of other habitats (e.g., rocky shores and bottoms, vegetation beds, over open sand in the case of Ehippidae) (Poyato-Ariza 2005). Fossils of pycnodont taxa such as Turbomesodon, Stenamara and Pycnodus also were found in freshwater and estuarine environments (Longbottom 1984; Poyato-Ariza & Wenz 2000; Poyato-Ariza & Wenz 2004).
The shape of the fins and their placement in several pycnodont specimens reveals that there is more morphological variation than initially assumed. The high number of more or less completely preserved specimens reveals that not all pycnodonts had long anal and dorsal fins. Genera such as Coccodus, Gibbodon and Ichthyoceras seemingly had rather short unpaired fins with few fin rays (ca. 6), whilst on the other extreme, Abdobalistum, Coelodus, Nursallia, and Oropycnodus had very elongated anal and dorsal fins with up to 60 fin rays (Poyato-Ariza & Wenz 2002). The caudal fin also varies in fin ray number from seven (Ichthyoceras) to nearly 40 (Abdobalistum). It is also extremely variable in shape from deeply forked to distal border convex.
More atypical pycnodonts are Gyrodus and Proscinetes. Their dorsal and anal fins are falcate and low, and are separated from the caudal fin by a deep caudal peduncle. The caudal fin is extremely deeply forked. The distribution of Gyrodus is very wide, ranging from Western Europe in the Oxfordian, Kimmeridgian and Tithonian to the western part of the Tethys (Kriwet & Schmitz 2005). It was also present in the western coast of what is now Chile during the Oxfordian and lived in what is now Siberia and Japan in the Tithonian (Kriwet & Schmitz 2005). This wide distribution and fin morphology suggests that both had pelagic lifestyles, swimming in open water close to the coast (Kriwet & Schmitz 2005).
Pycnodonts possess powerful molariform teeth indicating a durophagous diet (Nursall 1996). Even though they are interpreted as specialised shell crushers, a further look at the function and form of their jaws reveal that pycnodonts might be more flexible in their ecology. Kriwet (2001b) showed that while the premaxilla and maxilla are firmly fixed in basal pycnodonts, these bones become free and movable in more derived forms enabling protrusion of the upper jaws. Compared to teleosts, the protrusion seen in pycnodonts is quite limited as the premaxilla and maxilla connection is not as highly developed. Expansion of the bucco-pharyngeal cavity as indicated by reduced cheekbones in more derived taxa (Kriwet 2001b) might have assisted in suction feeding. This suction feeding, along with nipping and biting meant that pycnodonts as a group seemed to be more generalised benthic foragers with a wide range of prey (Poyato-Ariza 2005). Pycnodonts also have either incisiform or styliform teeth on the premaxilla and dentary, which also expanded the ecological functions within the group such as biting, gripping, grasping, scraping, picking, grazing, rasping and clipping (Kriwet 2001b). This combination of premaxillary/dentary dentition and the molariform dentition within the jaws indicates that pycnodonts could have ranged from generalised omnivores (Turbomesodon) to specialists (Mesturus) (Poyato-Ariza 2005).
A dental microwear analysis was performed on the teeth of several pycnodont genera (Gyrodus, Athrodon, Eomesodon, Proscinetes, Pycnodus) to more accurately determine their diet (Darras 2012). The results showed that durophagous pycnodonts are more similar to the generalised living durophage Archosargus probatocephalus than to the heavily specialised durophage Anarchias lupus. This shows that shellfish were just one of many prey items that these taxa would have fed on. Another result revealed that some pycnodonts might have been heavy grazers on algae. The combination of anterior teeth for cutting or excavating food and a mill within the jaws for processing algae is an efficient way to extract nutrients from tough food items. Such a structure is quite similar to the beak and pharyngeal mill present in parrotfishes (Labridae). A dental microwear analysis performed on parrotfishes could give more insight into how functionally similar pycnodonts were to these ecologically important herbivores. Possible herbivory in pycnodonts (Kriwet 2005; Darras 2012) alongside the recent discovery of multidentulate teeth in the Triassic semionotiform Hemicalypterus weiri (Gibson 2015) is compelling evidence against the hypothesis of fish herbivory arising in the Cenozoic (Bellwood 2003).
A study about the evolution of fishes on coral reefs from the Late Triassic to recent times showed that pycnodonts seemingly occupied significant morphospaces long before modern teleosts achieved the same after the K/Pg extinction (Goatley et al. 2010). Whilst teleosts with large eyes were common throughout the Cenozoic, pycnodonts have had taxa with relatively large eyes since the Late Triassic. In modern fishes, large eyes and large mouths indicate specialisation for nocturnal feeding. With their relatively large jaws combined with large eyes, it appears likely that at least some members of the pycnodonts could have been capable of nocturnal or low-light feeding. Most nocturnal fishes today are rarely durophagous with the exception of grunts (Haemulidae) and emperors (Lethrinidae), which feed on heavily shelled molluscs and crustaceans in off reef sediments (Goatley et al. 2010). Nocturnal durophages that occur on reefs could be one of the few unique ecological niches that pycnodonts occupied.
A recently described family of pycnodonts, Gladiopycnodontidae, discovered in Lebanon showcases the remarkable range of morphologies within these fishes (Taverne & Capasso 2013). These fishes are fusiform in shape with an elongated snout, which forms a rostrum protruding above the lower jaw. Pectoral fins are absent and replaced by a spine. Many members of this family also possess frontal, occipital, nuchal and cleithral spines. Only few characters (presence of dermosupraoccipital, long parasphenoid, infraorbitals) identify these fishes as pycnodonts. Gladiopycnodontid fossils are found in rocks dating from the Cenomanian in the Late Cretaceous; a time when pycnodonts seemingly were at the peak of their diversity. It has been suggested that their extreme covering of spines was used as protection against predators (Marramà et al. 2016a). The bizarre morphology of gladiopynodontids, at first glance, appears to have no modern counterpart but Marramà et al. (2016a) argued that their armoured trunk and the counterbalance of the long anterior rostrum with the sword shaped posterior anal plate suggests that they might have swam vertically with the head pointed downwards like fishes of the family Centriscidae. These small fishes use their long tubular mouth to pick off zooplankton and small benthic organisms. It is probable that Gladiopycnodus at least, also swam and fed in such a manner. Nevertheless, the taxonomy and systematics of this group, which is placed into Coccodontoidea remains ambiguous.
Nursall & Capasso (2004) described a peculiar taxon, Gebrayelichthys, with a remarkable appearance from the Late Cretaceous (Cenomanian) of Lebanon, which has a laterally compressed and very deep (nearly twice as deep as it is long) body. A bony mast protrudes upwards from the posterior end of the skull to form a high, narrow dorsal crest supporting the basal elements of the dorsal fin. The ventral keel is stouter than the dorsal mast but extends downwards to an equal extent as the dorsal mast protrudes upward. No other fish fossil or recent has such a bizarre body shape and could be described as hypsistisomatic or the “highest-bodied” fishes (Nursall & Capasso 2004). Several characters support the pycnodont affinities of this taxon (Taverne & Capasso 2014). A second taxon, Maraldichthys Taverne & Capasso, 2014, is placed into the family Gebrayelichthyidae alongside Gebrayelichthys. The morphology of gebrayelichthyids is such a radical departure from other pycnodonts that it is assumed that they were ecologically quite unique within Pycnodontomorpha (Nursall & Capasso 2004). They would have been slow swimming, mid water inhabitants of shallow seas. Trailing spines along the dorsal crest would have provided adequate defence against predators. Its teeth were sharp but few in number and it might have captured large plankton and slow moving nekton such as ctenophores, pteropods and larval fish.
A deeper look at pycnodonts reveals that they are far from the morphologically homogenous, ecological specialists that they appear to be at first sight. When looking at the surprising variation in their morphology, the true range of their distribution into a wide variety of habitats and their ecologically versatile mixture of generalists and specialists then it is of little surprise that their reign lasted for 175 million years.
|
up |
|
|
3.Goals |
The major goals of this project are to gain deeper insights and a better understanding of the mechanisms that determine the evolutionary history, success, and final extinction of a highly diverse clade of fishes, the Pycnodontiformes. It was demonstrated that pycnodont fishes were very successful, highly diversified and evidently well adapted to the habitats they occupied. It is hypothesized that they were an important and major element of marine as well as continental influenced fish faunas (e.g., Kriwet 2001a). Nevertheless, the reasons for (1) their seemingly rapid diversification in the Early Mesozoic, (2) their subsequent success and thus their possible superiority over other fish groups inhabiting same environments (e.g., Ginglymodi), (3) their habitat use (= facies depending distribution) and possible size aggregations due to ontogenetic shifts, (4) diversity fluctuations in, i.e., relation to abiotic crises (e.g., Cenomanian/Turonian and K/P boundary events, environmental changes) or biotic factors (e.g., competition), and (5) the events triggering their final disappearance more or less simultaneously with the appearance and evolution of teleostean groups, which are considered to be important elements of modern coral-fish assemblages are ambiguous or have not been addressed with rigorous analytical methods up to now. The proposed project intends to find answers to these still not fully understood and varied aspects, which (in addition with already known traits such as feeding kinematics) will help understanding the evolutionary history of this important group and thus has the potential to identify universal evolutionary processes.
|
up |
|
|
4.Specific subjects |
The project thus focuses on three major topics:
(1) Determination if there is a correlation between diversification patterns and biotic factors, particularly competition or abiotic factors,
(2) How pycnodonts were able to occupy their niches despite competition from ginglymodian fishes throughout the Mesozoic,
(3) Wether competition from teleosts in the Eocene was a factor in the extinction of this clade of fishes. To answer these questions, a multidisciplinary approach including different palaeobiological approaches is necessary.
|
up |
|
|
5. Innovative aspects, expected outcomes, and intellectual merit |
This project has the potential to significantly augment our understanding of macro- and co-evolutionary patterns in fishes through time by integrating different aspects. We expect that the results of this project will significantly advance and inform how certain clades prevail over others and give insights into the evolutionary success, ecology and extinction of pycnodonts but also other, related groups. The effects of abiotic factors on the diversification and extinction of clades is well studied but not for biotic factors such as competition. Innovative aspects of this project are the integrated approaches and analytical procedures, combining up-to-date fossil record, comparative phylogenetic, evolutionary, and morphospace analyses. Thus, to accomplish the main goals of the proposed project, our approach is a multidisciplinary one. All data acquired will be made available to the community for further scrutiny and research.
While morphospace analyses have been performed before on pycnodonts, it has never been done in conjunction with ginglymodians to track their changes in morphospace occupation through geological time. This will enable us to identify possible patterns such as the extent of morphospace overlap between the clades and what this reveals about the effects of interaction of species on clades that seem to share similar ecomorphologies. Marine ginglymodians have also never been used in morphospace analyses before and may lead to further inquiries into their marine ecology, which is insufficiently studied. The rise of teleosts, particularly, acanthomorphs has also been suggested as one of the primary reasons for the extinction of pycnodonts but has never truly been tested. Disparity and morphospace analyses between pycnodonts and teleosts of the Eocene might show how similar/different the morphospace occupation was and could be a useful starting point in truly looking at the demise of pycnodonts. Also, looking at the extent of morphospace occupation between different size classes of certain pycnodont genera could reveal how pycnodonts partition resources amongst conspecifics in the same habitat or if they occupied separate habitats to avoid competition. This information is crucial to understand and distinguish displacement and extinction patterns not only in extinct but also extant fishes related to biotic (competition, fisheries) and/or abiotic (e.g., climatic) factors. In this, the project represents not only pure research but also has an applied perspective, which can be used by conservation biologists to identify causes underlying diversity fluctuations and range displacements in extant fishes.
|
up |
|
|
6. Results |
Although the support by the FWF ended in May 2021, the project continues as many data that were collected during this project still need to be analysed, such as freshwater occurrences of pycnodonts, taxonomic identities of many still undescribed specimens, revisions of already described taxa, and their inter- and intrarelationships. Therefore, the results presented here are sort of preliminary and will be updated when necessary.
|
up |
|
|
6.1 Taxonomy |
In the course of the project, three new taxa were identified. All three are from the Late Cretaceous, (Cenomanian-Turonian), a time span when pycnodont diversity was at its highest (Marramà et al. 2016). Additionally, they all belong to the family Pycnodontidae, which is among the most diverse family in the order Pycnodontiformes. The first of these new species is Scalacurvichthys naishi from the Cenomanian of Israel from a quarry near the village of Ein Yabrud (Cawley & Kriwet 2018). Its forward facing first dorsal ridge scale and a triangular dermatocranium are some of the many characters that diagnose this specimen as a new taxon. The second new taxon, Flagellipinna rhomboides, is a diamond shaped pycnodontid with an elongated first fin ray of the dorsal fin which hails from the extremely productive locality of Haqel, Lebanon of the Cenomanian (Cawley & Kriwet 2019). The third new taxon is the pycnodontid Njoerdichthys dyckerhoffi from the Turonian of the Lengerich localities of Northern Germany (Cawley et al. 2020). Of significant interest concerning this species is that it comes from black shales at a higher than normal latitude for pycnodonts from the Turonian of the Late Cretaceous when the Cretaceous Thermal Maxima (KTM) occurred (Huber et al. 2018), suggesting pycnodont distribution to be heavily influence by climatic changes.
The taxonomic status of one of the last representatives of the pycnodont fishes in the fossil record, the genus Pycnodus from the extremely rich Bolca Konservat-Lagerstätte of the Early Eocene, also was re-considered (Cawley et al. 2018). The famous Bolca locality has produced specimens of Pycnodus in great abundance, which makes it a suitable candidate for research questions regarding taxonomic matters. Pycnodus has had a long, complex taxonomic history (Volta 1796; Blainville 1818; Agassiz 1833-1844; Heckel 1856; Blot 1987; Poyato-Ariza and Wenz 2002; Taverne 2003; Poyato-Ariza 2013) and the Bolca Lagerstätte specimens had three specific names assigned to them; Pycnodus apodus, P. platessus and P. gibbus. While P. platessus unambiguously represents a junior synonym of P. apodus (Agassiz 1833; Poyato-Ariza and Wenz 2002), there has been more controversy surrounding P. gibbus. Heckel (1856) described P. gibbus as its own distinct species based on the ‘gibbosity’ on the forehead, number of interdigitations between vertebrae and differences concerning the ratio of body length to body depth and vertebrae length to body depth. Conversely, Agassiz (1833-1844) identified these specimens as juveniles of P. platessus. Blot (1987) compared the anatomy of specimens of P. platessus and P. gibbus and concluded that the specimens are synonymous with each other and the variation contained within these specimens was intraspecific in nature. To confirm if this conclusions presented by Agassiz and Blot are valid, quantitative methods in the form of geometric morphometrics on specimens of Pycnodus that have been labeled in museum collections as Pycnodus sp., P. apodus, P. platessus or P. gibbus were employed to test if such labeled specimens separate from each other to warrant the names assigned to them or if the variation is restricted to one species of Pycnodus. In the case of P. platessus, it was included to investigate whether it overlaps with P. apodus and to verify its current status as a junior synonym. It is obvious that much overlap was observed in the morphospace of different labelled Pycnodus ‘species’ revealing that different species cannot be distinguished in the specimens studied here using geometric morphometrics. The main difference between specimens concerned eye size and body depth with larger eyed, deeper bodied forms being much smaller than the small eyed, streamlined forms. Since larger eyes typically belong to juvenile fishes (Pankhurst and Montgomery 1990), these deep-bodied forms are interpreted as juveniles of Pycnodus. The description of P. gibbus by Heckel (1856) also corresponds to the juvenile form with only meristic and morphometric characters distinguishing it from other specimens of Pycnodus. The results thus show P. gibbus to be an invalid species. Moreover, Pycnodus platessus has to be considered a synomym of P. apodus Morphometric characters and meristic counts also could not distinguish two or more different species of Pycnodus. Since apodus was the species name given to the holotype specimen of Pycnodus, all Bolca Lagerstätte specimens of this genus therefore have to be assigned to P. apodus. The clear difference between juvenile and adult forms also indicates niche partitioning between the two life history stages and we hypothesize that the juvenile would have keep close to the shelter of the reef while the adults would have been midwater dwellers that cruised over the reef.
|
up |
|
|
6.2 Sexual dimorphism |
Pycnodontiforms are unusual in that there are several recorded instances of sexual dimorphism known in this order, a phenomenon which is overall very rare to observe in the fossil record (Ostrom and Wellnhofer 1990; Sampson et al. 1997; Scannella and Fowler 2009). One form of sexual dimorphism in pycnodonts that can be observed is the difference in dorsal ridge scale morphology and number of spines on said scales in numerous specimens of Nursallia tethysensis (Capasso et al. 2009). Specimens of the gladiopycnodontid Gladiopycnodus karami also display different numbers of dorsal scutes (very modified dorsal ridge scales) (Taverne and Capasso 2013). Paranursallia gutturosum from the Late Cretaceous of Vallecillo, Mexico, showcases sexual dimorphism in the form of the males having an extremely elongated dorsal fin, the first 14 rays being three times the length of the female (Giersch 2014; Stinnesbeck et al. 2019). The holotype of the Lebanese pycnodontid Akromystax tilmachiton has a deeper skull and reduced curvature in the anterior border of its body in comparison to other specimens which Poyato-Ariza and Wenz (2005) speculate to be a sign of possible sexual dimorphism. By far the most extravagant example of sexual dimorphism in not just the pycnodonts but among all fossil bony fishes is the extraordinary range of horns and spines present in specimens of Hensodon spinosus (Capasso et al. 2010). What is believed to be the males have a pair of prominent horns in front of the orbits and a broad posterior spine present on the cleithrum.
An unusual specimen of the small gladiopycnodontid Pankowskichthys libanicus (Taverne and Capasso 2014a) housed in the MNHN Paris has enough unique cranial characteristics that differ from the holotype specimen, but postcranial characters support that both specimens belong to the same species (Cawley & Kriwet 2017). We interpret this as strong evidence of sexual dimorphism in this species. The distinguishing characters of the purported male specimen described in this chapter include: nuchal horn orientation, spacing of posterior spines on the nuchal horn and presence of parietal horns.
This new occurrence in combination with previous described sexual dimorphisms indicates that this was a common pattern among pycnodontiforms.

| Fig. 4.Holotype specimen of Pankowskichthys libanicus IRSNB P9278 interpreted here to be the female. Photo modified from Taverne and Capasso (2014). (b) P. libanicus, MNHN HAK 1950 showing the pair of short horns above the orbit and which is interpreted to be the male. Arrows used to show differences in cranial structures between two morphotypes. Black arrows, orientation of nuchal horn; White arrows, spaces between spines on nuchal horn; Blue arrows, position and shape of orbitosphenoid; Red arrows, horns above orbit present on parietal; Green arrows, pointed ventral tips on prefrontal. Scale bar = 1 cm. [from Cawley & Kriwet (2017)].
|
up |
|
|
6.3 Evolutionary success and extinction |
From the Late Triassic onwards, durophagy has been a very successful feeding strategy for actinopterygians (Lombardo and Tintori 2005) and three important neopterygian clades arose during this time that took advantage of this abundant resource of shelled molluscs and echinoderms. One of these were the pycnodonts while the other two were Dapediiformes and Ginglymodi. What were the factors that led pycnodonts to survive alongside the other shell crushing clades? Ginglymodians were fusiform reef dwellers that generally had powerful jaws with large molariform teeth that could crush the shells of mollusks, echinoderms and crustaceans (Jain 1984; Tintori 1996). Dapediiforms were more morphologically similar to pycnodonts due to their deep, rhomboid bodies with crushing jaws (Tintori 1983; Smithwick 2015). All three groups lived alongside each other during the Mesozoic with ginglymodians arising in the Middle Triassic (López-Arbarello et al. 2016) and both dapediiforms and pycnodonts making their first appearance in the Late Triassic (Tintori 1981, 1983). Ginglymodi were present in marine waters until the Early Cretaceous (Cavin 2010) where most forms moved into freshwater and this is where they have stayed until the modern day where there is only one surviving family of this once diverse clade, Lepisosteidae (gars). While dapediiforms also had freshwater representatives, they had most of their diversity in marine waters during their reign (Late Triassic-Late Jurassic) (Tintori 1983; Maxwell and López-Arbarello 2018). Throughout the Mesozoic and in the Palaeogene, pycnodonts were predominantly associated with shallow marine reef ecosystems.
While Ginglymodians and dapediiforms were common fish groups that lived alongside pycnodonts until the Early Cretaceous, in the Late Cretaceous another fish clade started to diversify and were common in shallow marine environments alongside pycnodonts: Acanthomorpha, the spiny finned teleosts. Today, these fishes dominate reef environments worldwide alongside most marine habitats and constitute a significant amount of freshwater fish diversity. It has been hypothesized that these fishes with their refined locomotory (Dewar and Graham 1994) and food handling adaptations (Liem, 1973; Wainwright 1989; Holzman et al. 2007, 2008a, b; Mabuchi et al. 2007; Wainwright et al. 2012, 2015; McGee et al. 2015) might have competed with pycnodonts to such a degree that they could have contributed to their extinction (Poyato-Ariza 2005a). However, a recent analysis of acanthomorph diversification in the aftermath of the K/Pg boundary event (Ghezelayagh et al. in review) revealed that the disparity observed in early Cenozoic fossil acanthomorphs most likely was the product of an extended period of phenotypic diversification demonstrating that the rise of acanthomorph diversity in the early Eocene was not punctuated, but a step-like process. Thus, the question why pycnodonts became extinct at the end of the Eocene remains a mystery.
Pycnodonts separate from other major durophageous Mesozoic groups, the ginglymodians and dapediiforms, in both body and jaw morphospace analyses (Cawley et al. 2021). The main difference in jaw morphology is the presence of a posterior coronoid process in pycnodont jaws, which indicate increased bite force and prey processing ability. Bite force is increased due to the vertical suspensorium present in many pycnodonts allowing adductor mandibulae muscles to attach in a more vertical orientation (Kriwet 2001c) and decreased jaw length (Gosline 1965). Prey processing is enhanced as the posterior position of the high coronoid process enable oral mastication (Ostrom 1966; Hoshi 1971; Erickson et al. 2012). This could give pycnodonts an advantage as they could extract more energy from prey as they could process tougher prey items than ginglymodians and dapediiforms and they would not need to spend as much energy digesting prey. While the two other groups would have had high coronoid processes which indicate high bite forces their anterior position would not allow them to mechanically process prey before digestion to the degree that pycnodonts could.
Body shape in pycnodonts is typically laterally compressed and deep bodied while dapediiforms are significantly less deep bodied particularly in Jurassic forms, which suggests they become more adapted to open water as the Mesozoic continues (Maxwell and López-Arbarello 2018). Ginglymodians are more fusiform but still bulky which supports the evidence that many were large bottom dwellers hovering over the sea bed (Tintori 1996). These morphometric differences in body and lower jaw suggest minimal competition between pycnodonts and other durophagous fish clades (Fig. 5). Additionally, there are significant differences between pycnodont families with Pycnodontidae comprised of deep bodied reef dwellers with powerful crushing jaws and gladiopycnodontids with more fusiform bodies and weakly developed jaws suggesting these were found over open sea bottoms preying on softer bodied invertebrates. Disparity results also show that jaw shapes are more disparate in pycnodonts than body shape indicating that niche partitioning in terms of targeting different prey would have been common in pycnodonts and could be a large reason why they were so successful.
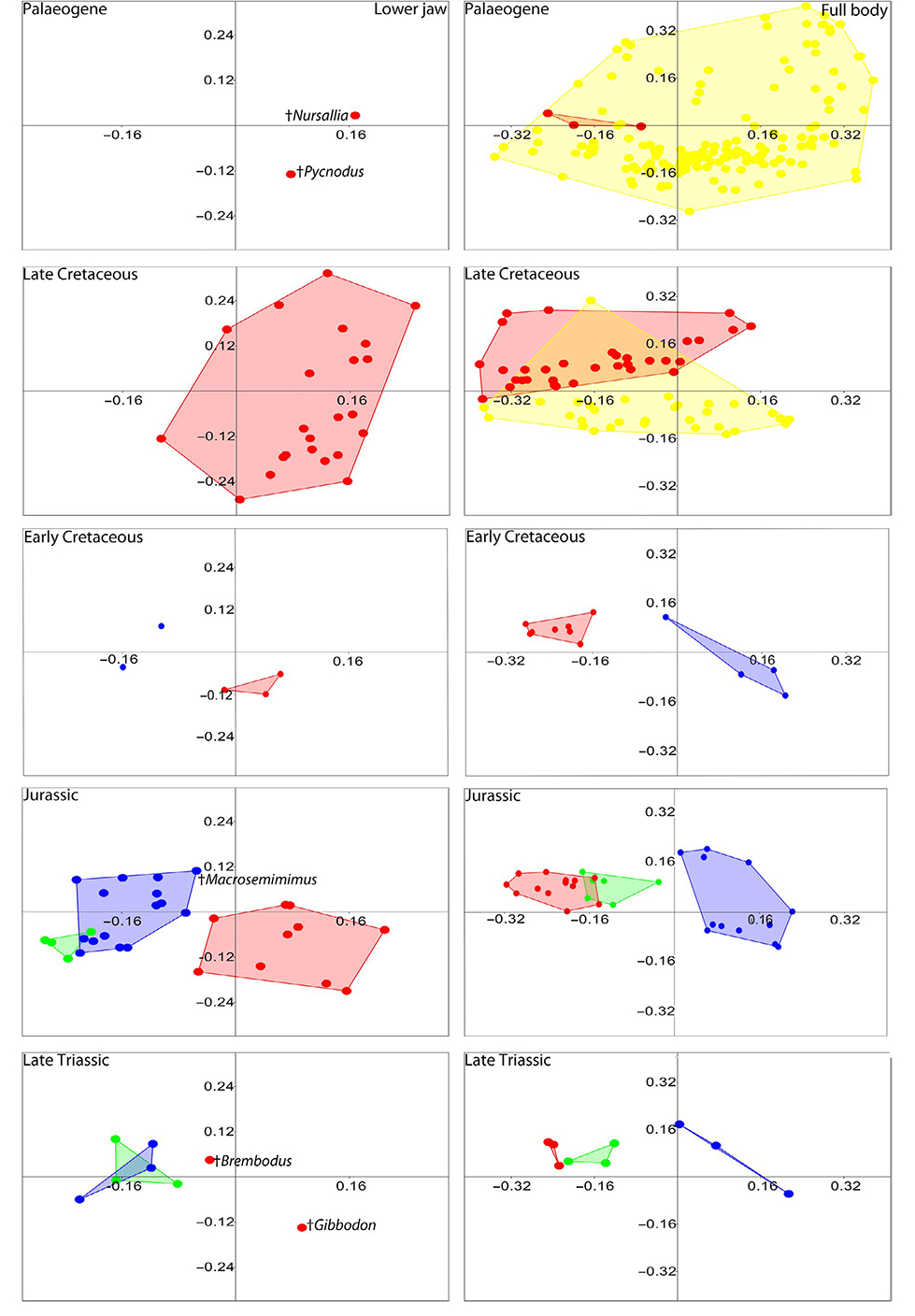
| Fig. 5.Morphospace occupation of pycnodontiforms (red), ginglymodians (blue), dapediiforms (green) and acanthomorphs (yellow) through time. Left: lower jaws. Right: full body shape. [from Cawley et al. (2020)].
In the Late Cretaceous, pycnodonts are separated from acanthomorphs to a significant degree. Things change in the Palaeogene where the species poor pycnodonts are completely overlapped by the far more speciose acanthomorphs, which occupy every quadrant of the morphospace. Since acanthomorph jaws are structured significantly differently that they cannot be directly compared to pycnodont jaws like the Mesozoic clades, we instead looked at the acanthomorph taxa that overlap with pycnodonts to see if they could have competed in prey acquisition. What we found were taxa that were either suction feeders on fishes and cephalopods and benthic generalists that would have fed on softer prey items. This along with the observation that durophagy (Santini et al. 2014) does not become a common mode of feeding in acanthomorphs until the Oligocene after the pycnodonts go extinct suggest that these fishes did not directly compete with these fishes so as to drive them to extinction. Indeed, the dominance of pycnodonts in the deep-bodied quadrant of the Late Cretaceous morphospace suggests that pycnodonts were reef specialists that could have kept acanthomorphs out of reef habitats and it was only after the K/Pg extinction that acanthomorphs colonized reef habitats.
Other than the Cenomanian-Turonian of the Late Cretaceous there was no correlation between diversity and sea level/sea surface temperature (SST). Indeed, the high diversity of the Late Jurassic pycnodonts is a time of lowered SST and sea level. We also looked at the invertebrate diversity present during the Palaeogene when the Palaeocene-Eocene Thermal Maxim (PETM) occurred which was an extinction event where SST was the highest in the Cenozoic (Thomas et al. 2002). This would have led to an increase in ocean acidification (Penman et al. 2014) which could have negatively impacted the shelled invertebrates which were common prey for pycnodonts. However, no correlation between invertebrate diversity and SST was found. Thus far, environmental changes seem to have little effect on pycnodont diversity. This changes when one looks closely at the amount of reef area through time (Kiessling et al. 1999). Here, it can be clearly seen that the highest amounts of reef area correlate very well with pycnodont diversity and that prime pycnodont habitat such as rudist bivalve reefs were declining before the K/Pg extinction and finally went extinct at the K/Pg boundary. This also explains why pycnodont diversity was declining even before the K/Pg extinction event. While pycnodonts are present in other habitats such as freshwater and estuaries (Poyato-Ariza et al. 1998; Poyato-Ariza 2005b; Cavin et al. 2020) or neritic environments (Kriwet and Schmitz 2005) they are predominantly an order specialized for reef life and such a reduction in habitat would have seriously impacted their numbers. Acanthomorphs, conversely, were adapted to a far wider range of habitats and when reefs began to recover, colonized and diversified relatively quickly (Friedman 2010). Pycnodonts obviously were “a dead clade walking”, a process that started already before the K/Pg boundary event, until they eventually dwindled to the point of being a victim of background extinction.
Conclusions (from Cawley et al. 2021)
- The majority of pycnodontiforms were specialized for manoeuvrability in reef-like environments with only few forms adapted also to open water habitats (e.g., †Gyrodontidae) and had different jaw structures, which avoided any potential competition with dapediiforms and ginglymodians.
- While both dapediiforms and ginglymodians overlap in jaw morphology their different body shapes indicate that they most likely occupied different niches or microhabitats thereby excluding any possible competition.
- The differences between pycnodontiforms and other neopterygian lineages including durophagous forms could be related to improved jaw performance for feeding on tougher organisms. Altered feeding mechanisms such as mastication seemingly were improved in pycnodontiforms for prey processing, while the ontogenetic increase of the size of the fish allowed access to differ- ent types of prey and/or specialize on one or a few species of prey.
- Pycnodontiform families also separate from each other in both the jaw and body morphospaces showing that they were most likely more diverse in their diets and habitat occupation than traditionally assumed. Gladiopycnodontids represent a significant expansion of pycnodontiform jaw morphospace, which is interpreted here as them occupying a more open, demersal habitat with new types of prey requiring different jaw morphologies and related soft structures such as muscles and ligaments. Hence the elongate, dorso-ventral compact jaws most likely were used for picking small benthic prey off the substrate. Our analyses show
that pycnodontiforms reduced competition with similar duropha- gous lineages by modifying the structures of the jaws for mastication of prey and further modification allowed pycnodontiforms to target different prey. While in larger taxonomic units, the difference in disparity is lower (Pycnodontiformes, Ginglymodi) the differences are larger between families, sometimes even signifi- cantly (Dapediidae, Brembodontidae, “Trewavasiidae”), which is indicative of further niche partitioning.
- Comparing the diversity patterns of different Mesozoic fish groups also reveals that pycnodontiforms were not negatively or positively affected by diversity patterns in other clades, further indicating that competition between these groups was minimal or even absent. Competition with acanthomorphs in terms of body shape was minimal in the Late Cretaceous with pycnodontiforms restricted to more structured habitats, while acanthomorphs in- habited the biotopes between and outside such structures.
- By the Palaeogene, acanthomorphs had significantly expanded their morphological disparity to the point that many repre- sentatives shared the same body shape with pycnodontiforms. However, competition with pycnodontiforms in terms of feed- ing ecology was highly unlikely, as teleosts do not show a genuine radiation of extreme durophagous forms until the Oligocene after the Eocene climatic optimum event and after pycnodontiforms went extinct. Consequently, we rule competition with acanthomorphs as the reason for pycnodontiform extinction out. Conversely, morphospace results show that pycnodontiforms may have kept acanthomorphs out of reef habitats and it was due to habitat loss in the Late Cretaceous that acanthomorphs experienced their rapid speciation during the Palaeogene. With pycnodontiform diversity already in decline, new niches were open for these more recent neopterygians to fill and we hypothesize that when acanthomorphs started to dominate reef fish communities, pycnodontiforms were effectively a “dead clade walking”, becoming a victim to background extinction rather than any significant environmental changes or absence of possible prey. Rather than pycnodontiforms being outcompeted by the more derived teleosts, it appears that teleostean fishes only took over reefs after pycnodontiforms were already beginning to decline and only developed durophagous forms after their final extinction.
- The decline of reefs, particularly the extinction of rudist reefs, during the Late Cretaceous could be a promising avenue for future research in regards to abiotic and biotic drivers of pycnodontiform decline and extinction.
|
up |
|
|
6.4 Intrarelationships, compltness of fossil specimens and fossil record |
This part of the project is still ongoing and only some preliminary results are presented here including a first, tentative (new) phylogenetic scheme.
Despite the renewed awareness that pycnodonts have received in recent years in terms of phylogeny (Fig. 6), there are several gaps in our knowledge of their relationships. One of the largest relates to the discovery of two new families since the work of Poyato-Ariza and Wenz (2002): Gebrayelichthyidae and Gladiopycnodontidae both including highly derived forms from the Lebanese limestones of the early Late Cretaceous (Nursall and Capasso 2004; Taverne and Capasso 2013). These fishes differ significantly from the typical body form of pycnodonts with extreme body shortening and body depth in gebrayelichthyids and the fusiform armoured bodies of gladiopycnodontids. One of the primary reasons for the phylogenetic analysis is to identify the systematic relationships of these bizarre fishes within Pycnodontiformes as no phylogenetic work has ever been done on these groups. Taverne and Capasso (2013) did suggest that Gladiopycnodontidae and Coccodontidae were allied together due to sharing twelve characters between them under the superfamily Coccodontoidea. Gebrayelichthyidae was also included in Coccodontoidea but still with no cladistic analysis performed to actually test these relationships (Taverne and Capasso 2014b). In order to understand diversification patterns in pycnodonts and to ascertain the origin of certain morphological characters within the group, a newly updated phylogeny will serve as a vital framework to help researchers ask such questions. Moreover, the influence of specimen completeness on the topology also was tested.
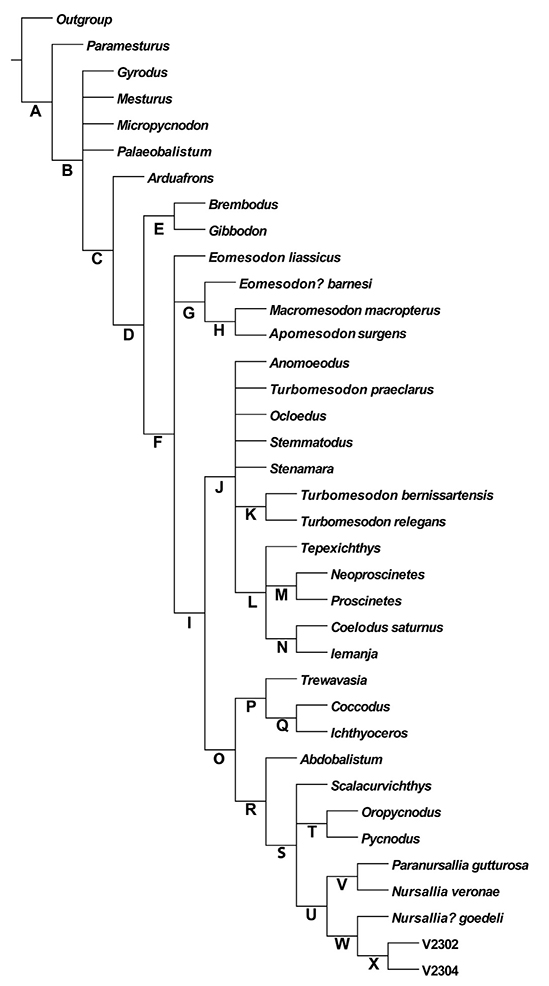
| Fig. 6.Most recent phylogenetic hypothesis of pycnodontiform relationships from Cawley et al. (2020) based on a data matrix of 37 taxa and 88 characters. All characters treated unordered and unweighted.
By breaking down qualitative characters from authors such as Poyato-Ariza and Wenz (2002) and Ebert (2020) that originally had three to four states into just having a binary state and including 156 new characters it confirms that the more characters that are analysed in a phylogenetic analysis, the higher the resolution of the tree will be (Prevosti and Chemisquy 2010; Puttick et al. 2019). The inclusion of derived families such as Gladiopycnodontidae and Gebrayelichthyidae revealed that both families are monophyletic (except for Ichthyoceros being outside of Gladiopycnodontidae). Additionally, certain clades such as the subfamily Pycnodontinae continue to be monophyletic and matches similar results of Poyato-Ariza (2020). Clades that were derived in previous analyses such as Nursallinae (Poyato-Ariza and Wenz 2002; Ebert 2020; Poyato-Ariza 2020) are found in more basal positions here even being found outside the family Pycnodontidae altogether. This could be due to the taxa within that subfamily having a comparatively high number of missing characters with the most basal taxon Abdobalistum being the most incomplete of them all with 150 characters missing out of a total of 424 (64.62% completeness). As missing characters can move taxa towards the stem of the tree (Sansom 2015) I assume that this is the case for Abdobalistum and its sister taxa. Interestingly, the consensus tree produced has high levels of homoplasy yet the tree was well resolved with very little polytomy. Another intriguing finding is that removing uninformative characters from the analysis did not improve the Consistency index (CI) or Retention Index (RI) and even reduced the resolution. Another encouraging aspect of this result was achieved by a straightforward Parsimony Analysis and more refined methods should hopefully elucidate the relationships between pycnodont taxa further. Problematic taxa such as Abdobalistum and Stenoprotome, which comparatively have a high number of missing characters, might reduce the homoplasy of the strict consensus tree if they are removed from the analysis. However, both of these taxa are above the standard for what are considered highly incomplete taxa (0-25% complete). Research has also shown that incomplete taxa are not detrimental to the accuracy of phylogenetic trees due to preventing long-branch attraction and preserving combinations of characters not found anywhere else (Wiens 2003, 2005, 2006). Additionally, even incomplete taxa can be accurately placed if the number of characters is high which is the case in my analysis performed here. The phylogeny produced here is confirmation of many of the simulated findings produced by authors such as Wiens (2003, 2006), Prevosti and Chemisquy (2010) and Puttick et al. (2019).
The results that have been obtained so far produced an intriguing consensus tree as while it had a low Consistency Index (CI) and Retention Index (RI) (something, which is to be expected as more characters produce more homoplasy increasing the number of tree steps (Al Sayad and Yassin 2019)), the resulting tree had a rather high resolution with only nine nodes collapsed. This result supports the work of authors such as de Carvalho (1996), Källersjö et al. (1999), and Radel et al. (2013), who found that homoplasy can improve the resolution of relationships in a phylogenetic tree. Further analysis will include the Bayesian MK model as this method is currently identified as the best method for recovering the most accurate trees when homoplasy is as high as the results produced here (Puttick et al. 2019).
|
up |
|
|
7. Publications |
In prep.
- Cawley, J.J. & Kriwet, J. in prep. Homoplasy and completeness of fossil specimens in phylogenetic relationships and how it might increase understanding of pycnodont evolution.
2021
- Cawley, J.J., Marramà, G., Carnevale, G., Villafaña, J.A., López-Romero, F., Kriwet, J., 2021. The rise and fall of †Pycnodontiformes: Diversity, competition and extinction of a successful fish clade. Ecology and Evolution, 11: 1769-1796. Link to article
2020
- Cawley, J.J., Lehmann, J., Wiese, F. & Kriwet, J. 2020. Some like it hot: Njoerdichthys dyckerhoffi gen. et sp. nov. (Pycnodontiformes, Lower Turonian) northward migration caused by the Cretaceous Thermal Maximum. Cretaceous Research, 116: 104590. Link to article
2019
- Cawley, J.J. & Kriwet, J. 2019. A new pycnodontid fish Flagellipinna rhomboides (Neopterygii, Pycnodontiformes) from the Upper Cretaceous (Cenomanian) of Lebanon, with notes on juvenile form and ecology. Journal of Vertebrate Paleontology, 39: e1614012 (16 pages). Link to article
- Gouiric-Cavalli, S., Remírez, M. & Kriwet, J. 2019. New pycnodontiform fishes (Actinopterygii, Neopterygii) from the Early Cretaceous of the Argentinian Patagonia. Cretaceous Research, 94:45-58. Link to article
2018
- Cawley, J.J., Marramà, G., Carnevale, G. & Kriwet, J. 2018. A quantitative approach to determine the taxonomic identity and ontogeny of the pycnodontiform fish Pycnodus (Neopterygii, Actinopterygii) from the Eocene of Bolca Lagerstätte, Italy. PeerJ, 6:e4809 (30 pages). Link to article
2017
- Cawley, J.J. & Kriwet, J. 2017. A new pycnodont fish, Scalacurvichthys naishii, gen. et sp. nov., from the Late Cretaceous of the Middle East. Journal of Systematic Palaeontology, 16: 659-673. Link to article
- Cawley, J.J. & Kriwet, J. 2017. Possible sexual dimorphism in Pankowskichthys (Neopterygii, Pycnodontiformes) from the Cenomanian, Haqel, Lebanon. Research & Knowledge 3: 33-35. Link to article
- Cawley, J.J. & Kriwet, J. 2017. New information about Late Cretaceous pycnodont fishes (Actinopterygii, Pycnodontiformes) from the Near East. Research & Knowledge 3: 47-48. Link to article
- Stumpf, S., Ansorge, J., Pfaff, C. & Kriwet, J. 2017. Early Jurassic diversification of pycnodontiform fishes (Actinopterygii, Neopterygii) after the end-Triassic extinction event: Evidence from a new genus and species, Grimmenodon aureum. Journal of Vertebrate Paleontology 37(4): e1344679 (14 pages). Link to article
|
up |
|
|
8. Conference abstracts |
2019
- Cawley, J.J., Marramà, G., Carnevale, G., Kriwet, J. 2019. Investigation of the evolutionary success and extinction of pycnodont fishes using quantitative methods. Abstracts of the 79th Annual Meeting of the Society of Vertebrate Paleontology: 76.
- Kriwet, J., Marramà, G., Carnevale, G. & Cawley, J.J. 2019. Success and demise of pycnodont fishes (Neopterygii, †Pycnodontiformes). – In: Nützel, A., Reichenbacher, B. & Krings, M. (eds): Paleo & Life – Abstracts of the 90th Annual Meeting of the Paläontologische Gesellschaft, Munich: 85; SNSB-BSPG, München.
- Kriwet J., Marramà, G., Carnevale, G. & Cawley, J.J. 2019. Drivers of taxonomic and functional diversity in extinct non-teleostean neopterygians (Osteichthyes, Actinopterygii). Journal of Morphology, Supplement 280 (International Congress of Vertebrate Morphology (ICVM) Abstract Issue): 156.
2018
- Cawley, J.J. & Kriwet, J. 2018. Pycnodonts: Could they be the true sister group to teleost fishes?. – In: Schwarz, C., Zimmermann, D. & Kriwet, J. (eds).19. Jahrestagung Gesellschaft für Biologische Systematik, Abstract volume: 12; Verlag Dr. Friedrich Pfeil, Munich.
- Kriwet, J., Marramà, G. & Cawley, J.J. 2018. Predicting competition between †Pycnodontiformes and Ginglymodi (Osteichthyes, Neopterygii) through geologic time based on quantitative analyses of lower jaw features. 5th International Paleontological Congress, Abstract Volume: 440, Paris.
|
up |
|
|
9. Conference participations |
2019
- 79th Annual Meeting of Society of Vertebrate Paleontology
Brisbane, Australia (Oct. 9-12)
Talk: Investigation of the evolutionary success and extinction of pycnodont fishes using quantitative methods (co-authored by Marramà, G., Carnevale, G. & Kriwet, J.).
- 90th Annual Meeting of the Paläontologische Gesellschaft
Munich, Germany (Sep. 15-18)
Talk: Success and demise of pycnodont fishes (Neopterygii, †Pycnodontiformes) (co-authored by Kriwet, J., Marramà, G. & Carnevale, G.).
- AK Wirbeltierpaläontologie (Paläontologische Gesellschaft)
Vienna, Austria (March 15-17)
Poster: A quantitative approach to determine the taxonomic identity and ontogeny of the pycnodontiform fish Pycnodus (Neopterygii, Actinopterygii) from the Eocene of Bolca Lagerstätte, Italy (co-authored by Marramà, G., Carnevale, G. & Kriwet, J.).
- 12th International Congress of Vertebrate Morphology
Prague, Czech Republic (July 21-25)
Talk: Drivers of Taxonomic and Functional Diversity in Extinct Non-Teleostean Neopterygians (Osteichthyes, Actinopterygii) (co-authored by Kriwet, J., Marramà, G. & Carnevale, G.).
2018
- 9th Annual Meeting of the Gesellschaft für Biologische Systematik
Vienna, Austria (Feb. 11-14)
Talk: Pycnodonts: could they be the true sister group to teleost fishes? (co-authored by Kriwet, J.).
- 5th International Palaeontological Congress
Paris France (July 9-13)
Talk: Predicting competition between †Pycnodontiformes and Ginglymodi (Osteichthyes, Neopterygii) through geologic time based on quantitative analyses of lower jaw features (co-authored by Kriwet, J. & Marramà, G.).
2017
- 7th International Meeting on Mesozoic Fishes
Mahasarakham, Thailand (Aug. 1-7)
Talk: New information about Late Cretaceous pycnodont fishes (Actinoptergyii,Pycnodontiformes) from the Near East (co-authored by J. Kriwet).
Poster: Possible sexual dimorphism in Pankowskichthys libanicus (Neopterygii, pycnodontiformes) from the Cenomanian, Haqel, Lebanon (co-authored by Kriwet, J.).
|
up |
|
|
|
|
|
